

 RSS Feed (xml)
RSS Feed (xml)
Bovine Spongiform
Encephalopathy Tipe-H Atipikal
Bovine Spongiform Encephalopathy Tipe-H Atipikal pada Sapi yang Lahir setelah Larangan Pakan yang Diperkuat pada Meat-and-Bone Meal di Eropa
RINGKASAN
Pentingnya bovine spongiform encephalopathies (BSE) atipikal pada sapi untuk mengendalikan epidemi BSE kurang dipahami. Di sini kami melaporkan kasus BSE tipe-H atipikal pada sapi yang lahir setelah penerapan larangan pakan yang diperkuat di Eropa. Ini mendukung etiologi BSE tipe-H yang tidak terkait dengan BSE klasik.
LAPORAN KASUS
Seekor sapi Red Angus berusia 6,5 tahun mengalami downer cow syndrome setelah kelahiran anak sapi yang mati di Swiss pada Februari 2012. Hewan tersebut lahir di Jerman pada tahun 2005 dan diimpor ke Swiss pada usia 17 bulan. Menurut pemiliknya, tidak ada tanda-tanda penyakit sapi sebelum melahirkan. Setelah penyembelihan darurat, sampel medula oblongata diambil sesuai dengan pengawasan hukum Swiss bovine spongiform encephalopathy (BSE). Tes cepat BSE awal (Check Western; Prionics) (21) yang dilakukan oleh laboratorium regional hasilnya positif. Akibatnya, sampel medula oblongata dikirim, bersama dengan otak yang tersisa, yang masih tersedia di rumah jagal, ke Laboratorium Referensi BSE Swiss. Di sana, hewan tersebut dikonfirmasi BSE positif dengan TeSeE Western blot (Bio-Rad) (2), menggunakan pencernaan proteinase K terbatas dan deteksi imun dengan dua antibodi monoklonal spesifik protein prion (MAbs), Sha31 (11) dan 12B2 (16) . Massa molekul proteinase K-resistant prion protein peptides (PrPres) di Western blot ditentukan dengan perangkat lunak Quantity One versi 4.6.2 (Bio-Rad). Dibandingkan dengan sampel kontrol BSE klasik (tipe-C), pita PrPres yang terlihat dalam kasus ini menunjukkan ∼1,3- hingga 1,4-kDa massa molekul yang lebih tinggi serta pita tambahan pada ∼7,2 kDa. Juga, sampel bereaksi dengan MAb 12B2 (Gambar 1). Hal ini konsisten dengan fenotip molekuler BSE tipe-H (14). Distribusi protein prion terkait penyakit (PrPd) di seluruh otak ditentukan dengan uji imunosorben terkait-enzim (ELISA) (kit uji antigen BSE-scrapie; Idexx). PrPd terdeteksi terutama di thalamus dan obex dan, pada tingkat lebih rendah, di korteks serebelar, hippocampus, lobus pyriformis, dan inti basal (Gambar 2). Analisis histopatologis dilakukan pada bagian parafin yang diwarnai hematoxylin-and-eosin (H&E) dari daerah otak yang sama dengan yang dianalisis dalam ELISA. Lesi spongiform minimal terdapat di daerah obex (Gambar 3a) dan di otak tengah, tetapi tidak di struktur otak lainnya. Dengan imunohistokimia (menggunakan MAb F99) (17), endapan PrPd ringan diamati pada nukleus motorik dorsal saraf vagus, nukleus olivary kaudal (Gambar 3b), nukleus cuneate (Gambar 3c), nukleus hipoglosal, nukleus traktus spinalis saraf trigeminal, dan nukleus traktus soliter (Gambar 3d), serta di otak tengah dan thalamus. Endapan ini bertipe partikulat kasar, intraneuronal, dan intraglial. Tidak ada pelabelan PrPd di otak kecil, hippocampus, inti basal, dan korteks serebral. Seluruh kerangka bacaan terbuka dari protein prion sapi diurutkan dan tidak menunjukkan varian DNA dibandingkan dengan urutan referensi (aksesi GenBank no. AJ298878.1). Khususnya mutasi E211K yang dianggap menyebabkan varian genetik BSE tipe-H (19) tidak ada. Setelah konfirmasi laboratorium penyakit, bangkai hewan, termasuk semua produk sampingan, dimusnahkan, dan tidak ada bahan yang masuk ke rantai makanan.
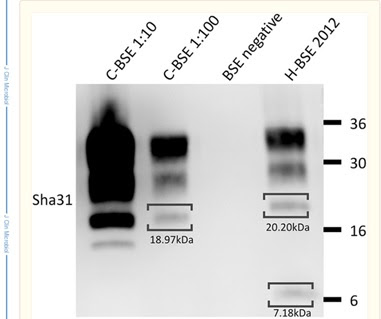

Gambar 1. Bio-Rad TeSeE hybrid Western blot menggunakan MAb Sha31 dan MAb 12B2. Massa molekuler dari peptida protein prion (PrPres) tahan proteinase K individu ditunjukkan di bawah tanda kurung. Perhatikan perbedaan dalam massa molekul dan reaktivitas 12B2 antara kasus BSE tipe-H dan kontrol BSE tipe-C serta PrPrespeptide tambahan pada ∼7 kDa.

BAGIAN OTAK
Gambar 2. Distribusi protein prion patologis neuroanatomi (PrPd). Distribusi PrPd di otak kasus BSE tipe H ditentukan dengan uji Idexx. Cutoff tes ditunjukkan oleh garis putus-putus.

Gambar 3. Histopatologi dan imunohistokimia. (a) Inti motorik dorsal saraf vagus (pewarnaan H&E). Vakuola ditunjukkan oleh panah. ( B ) Inti olivary kaudal dengan pelabelan PrPd intraneuronal dominan. ( c ) Cuneate nukleus, intraneural (panah), dan pelabelan PrPd intraglial dan pewarnaan neuropil yang ringan. Pola pewarnaan serupa diamati pada nukleus saluran tulang belakang trigeminus. ( d ) Inti saluran soliter dengan pelabelan PrPd glial jarang.
BSE adalah penyakit menular dan neurodegeneratif yang muncul di Inggris pada pertengahan 1980-an dan kemudian di benua Eropa, Jepang, dan Amerika Utara (26). Hal ini disebabkan oleh prion, yaitu protein prion seluler (PrPd) yang salah lipatan yang menumpuk di otak sapi yang terkena. Penyakit prion dapat diperoleh, (yaitu, ditularkan melalui infeksi), memiliki dasar genetik, atau berkembang secara spontan sebagai kasus sporadis (9). Tiga jenis BSE saat ini dibedakan: tipe C-, L-, dan H. Sementara BSE tipe C sejauh ini merupakan bentuk penyakit yang paling sering, BSE tipe L dan H, juga disebut sebagai BSE atipikal, adalah kondisi langka yang menunjukkan karakteristik yang berbeda secara biokimia dan biologis dari BSE tipe C (6, 8). BSE tipe-C diperoleh dan transmisi prion terjadi dengan menelan jaringan yang terinfeksi — pada ruminansia terutama tepung daging dan tulang (MBM) yang digunakan sebagai suplemen pakan (27). Karena masa inkubasi beberapa tahun, usia rata-rata sapi yang terkena BSE adalah 5 sampai 6 tahun selama epidemi, tetapi kasus juga telah diidentifikasi pada hewan yang jauh lebih muda dan lebih tua. BSE tipe-H hanya didiagnosis pada sekitar 30 sapi di seluruh dunia, semuanya berusia di atas 8 tahun (24). Deteksi sebagian besar dengan skema surveilans aktif (yaitu, dengan pengujian laboratorium terhadap sejumlah besar sapi dewasa yang tidak diduga menderita BSE klinis). Penentuan tipe strain biologis melalui transmisi eksperimental ke model hewan pengerat dan sapi menunjukkan bahwa BSE tipe-H disebabkan oleh agen prion yang berbeda dari BSE tipe C dan L (4, 5, 15, 18). Hingga saat ini, patologi dan etiologi BSE tipe-H masih kurang dipahami (24). Terutama pertanyaan apakah penyakit itu didapat, genetik, atau sporadis meninggalkan kesenjangan besar dalam pengetahuan kita. Kasus yang disajikan di sini memperluas pemahaman tentang BSE tipe-H dalam beberapa aspek.
Seluruh patologi otak dari BSE tipe-H hanya dilaporkan pada seekor banteng dari breed zebu mini (23, 25) dan pada sapi yang diinokulasi secara intraserebral secara eksperimental (15, 18). Untuk semua kasus BSE tipe-H alami lainnya, hanya sampel batang otak yang tersedia, dan tidak diketahui struktur otak mana yang merupakan target diagnostik yang paling cocok. Zebu berusia 19 tahun dan menunjukkan tanda-tanda neurologis yang menonjol, dan lesi spongiform serta endapan PrPd parah dan tersebar luas di otak. Sebaliknya, hewan yang dijelaskan di sini berusia 6,5 tahun, tanda-tanda neurologis khusus sistem saraf pusat (SSP) tidak diamati, dan lesi spongiform serta endapan PrPd di otak minimal. Semua temuan ini mendukung bahwa sapi tersebut berada pada tahap awal penyakit praklinis. Dalam hal ini, penting untuk menunjukkan bahwa lesi minimal dan endapan PrPd ini ditemukan pada struktur materi abu-abu di daerah obex medula oblongata, otak tengah, dan talamus. Temuan ini pada dasarnya mirip dengan BSE tipe-C praklinis (1, 12, 13, 22) dan mendukung bahwa pengambilan sampel wilayah obex dalam skema surveilans yang diterapkan untuk BSE tipe-C mungkin juga cocok untuk mendeteksi H- yang terjadi secara alami. ketik BSE.
Langkah pengendalian penyakit utama BSE tipe C adalah larangan MBM mamalia dalam pakan ruminansia. Larangan pakan ini diberlakukan di Swiss dan Uni Eropa pada awal 1990-an dan sangat mengurangi jumlah sapi yang baru terinfeksi. Namun, daur ulang agen BSE tipe C pada populasi sapi tidak diblokir sampai larangan pakan MBM diperkuat pada tahun 2001, sekarang tidak termasuk penggunaan protein hewani dalam pakan semua hewan ternak (10). Apakah BSE tipe-H juga ditransmisikan secara oral pada populasi sapi dengan MBM sebagai kendaraan masih belum diketahui. Jika transmisi oral terjadi dan merupakan satu-satunya etiologi, pelarangan pakan MBM yang diperkuat harus menjadi tindakan yang tepat untuk mencegah penyebaran BSE tipe-H juga, dan BSE tipe-H tidak boleh terdeteksi pada hewan yang lahir setelah penerapannya, yaitu, setelah tahun 2001 di Swiss dan Jerman. Sepengetahuan kami, ini adalah laporan pertama dari hewan yang terkena BSE tipe-H yang lahir setelah larangan pakan MBM yang diperkuat di negara masing-masing. Oleh karena itu, kasus ini memberikan bukti lebih lanjut bahwa etiologi BSE tipe-H mungkin tidak terkait dengan konsumsi daging dan tulang yang terkontaminasi prion. Secara bersama-sama, ini mendukung postulat yang diekspresikan secara luas bahwa BSE tipe-H berasal dari kesalahan lipatan spontan PrP seluler dengan patofisiologi yang mirip dengan penyakit Creutzfeld-Jakob sporadis pada manusia (7, 20). Sebagai alternatif, rute transmisi lain yang belum diketahui atau determinan genetik harus dipertimbangkan. Ini mengatakan, BSE tipe-H mungkin bertahan setelah pemberantasan BSE tipe-C. Apa implikasi dari skenario ini? Studi dengan tikus memberikan bukti eksperimental bahwa BSE tipe-H dapat mengubah fenotipe penyakitnya menjadi BSE tipe-C (3) setelah transmisi. Oleh karena itu telah dihipotesiskan bahwa epidemi BSE tipe-C berasal dari kasus BSE tipe-H yang terjadi secara spontan. Jika ini kasusnya, akan selalu ada risiko bahwa BSE tipe C akan muncul kembali pada populasi sapi setelah larangan pemberian pakan dihentikan. Akibatnya, beberapa tindakan pengendalian penyakit perlu dipertahankan tanpa batas waktu. Karena standar penentuan status risiko BSE suatu negara saat ini tidak membedakan antara subtipe BSE (28), penilaian risiko BSE tentu perlu mempertimbangkan pertimbangan tersebut. Ini menyoroti kebutuhan untuk melanjutkan penelitian tentang hubungan antara varian BSE klasik dan atipikal untuk memberikan dasar ilmiah untuk kebijakan pengawasan dan pengendalian penyakit di masa depan.
DAFTAR PUSTAKA
1. Arnold ME, et al. 2007. Estimating the temporal relationship between PrPSc detection and incubation period in experimental bovine spongiform encephalopathy of cattle. J. Gen. Virol. 88:3198–3208.
2. Arsac JN, Biacabe AG, Nicollo J, Bencsik A, Baron T. 2007. Biochemical identification of bovine spongiform encephalopathies in cattle. Acta Neuropathol. 114:509–516.
3. Baron T, et al. 2011. Emergence of classical BSE strain properties during serial passages of H-BSE in wild-type mice. PLoS One 6:e15839 doi:10.1371/journal.pone.0015839.
4. Baron TG, Biacabe AG, Bencsik A, Langeveld JP. 2006. Transmission of new bovine prion to mice. Emerg. Infect. Dis. 12:1125–1128.
5. Beringue V, et al. 2006. Isolation from cattle of a prion strain distinct from that causing bovine spongiform encephalopathy. PLoS Pathog. 2:e112 doi:10.1371/journal.ppat.0020112.
6. Biacabe AG, Laplanche JL, Ryder S, Baron T. 2004. Distinct molecular phenotypes in bovine prion diseases. EMBO Rep. 5:110–115.
7. Biacabe AG, Morignat E, Vulin J, Calavas D, Baron TG. 2008. Atypical bovine spongiform encephalopathies, France, 2001–2007. Emerg. Infect. Dis. 14:298–300.
8. Casalone C, et al. 2004. Identification of a second bovine amyloidotic spongiform encephalopathy: molecular similarities with sporadic Creutzfeldt-Jakob disease. Proc. Natl. Acad. Sci. U. S. A. 101:3065–3070.
9. Colby DW, Prusiner SB. 2011. Prions. Cold Spring Harbor Perspect. Biol. 3:a006833 doi:10.1101/cshperspect.a006833.
10. Ducrot C, Arnold M, de Koeijer A, Heim D, Calavas D. 2008. Review on the epidemiology and dynamics of BSE epidemics. Vet. Res. 39:15 doi:10.1051/vetres:2007053.
11. Feraudet C, et al. 2005. Screening of 145 anti-PrP monoclonal antibodies for their capacity to inhibit PrPSc replication in infected cells. J. Biol. Chem. 280:11247–11258.
12. Hoffmann C, et al. 2007. Prions spread via the autonomic nervous system from the gut to the central nervous system in cattle incubating bovine spongiform encephalopathy. J. Gen. Virol. 88:1048–1055.
13. Iwata N, et al. 2006. Distribution of PrP(Sc) in cattle with bovine spongiform encephalopathy slaughtered at abattoirs in Japan. Jpn. J. Infect. Dis. 59:100–107.
14. Jacobs JG, et al. 2007. Molecular discrimination of atypical bovine spongiform encephalopathy strains from a geographical region spanning a wide area in Europe. J. Clin. Microbiol. 45:1821–1829.
15. Konold T, et al. 2012. Experimental H-type and L-type bovine spongiform encephalopathy in cattle: observation of two clinical syndromes and diagnostic challenges. BMC Vet. Res. 8:22 doi:10.1186/1746-6148-8-22.
16. Langeveld JP, et al. 2006. Rapid and discriminatory diagnosis of scrapie and BSE in retro-pharyngeal lymph nodes of sheep. BMC Vet. Res. 2:19 doi:10.1186/1746-6148-2-19.
17. Nentwig A, et al. 2007. Diversity in neuroanatomical distribution of abnormal prion protein in atypical scrapie. PLoS Pathog. 3:e82 doi:10.1371/journal.ppat.0030082.
18. Okada H, et al. 2011. Experimental H-type bovine spongiform encephalopathy characterized by plaques and glial- and stellate-type prion protein deposits. Vet. Res. 42:79 doi:10.1186/1297-9716-42-79.
19. Richt JA, Hall SM. 2008. BSE case associated with prion protein gene mutation. PLoS Pathog. 4:e1000156 doi:10.1371/journal.ppat.1000156.
20. Sala C, et al. 2012. Individual factors associated with L- and H-type bovine spongiform encephalopathy in France. BMC Vet. Res. 8:74 doi:10.1186/1746-6148-8-74.
21. Schaller O, et al. 1999. Validation of a Western immunoblotting procedure for bovine PrP(Sc) detection and its use as a rapid surveillance method for the diagnosis of bovine spongiform encephalopathy (BSE). Acta Neuropathol. 98:437–443.
22. Schulz-Schaeffer WJ, Fatzer R, Vandevelde M, Kretzschmar HA. 2000. Detection of PrP(Sc) in subclinical BSE with the paraffin-embedded tissue (PET) blot. Arch. Virol. Suppl. 2000:173–180.
23. Seuberlich T, et al. 2006. Spongiform encephalopathy in a miniature zebu. Emerg. Infect. Dis. 12:1950–1953.
24. Seuberlich T, Heim D, Zurbriggen A. 2010. Atypical transmissible spongiform encephalopathies in ruminants: a challenge for disease surveillance and control. J. Vet. Diagn. Invest. 22:823–842.
25. Tester S, et al. 2009. Biochemical typing of pathological prion protein in aging cattle with BSE. Virol. J. 6:64 doi:10.1186/1743-422X-6-64.
26. Wells GA, et al. 1987. A novel progressive spongiform encephalopathy in cattle. Vet. Rec. 121:419–420.
27. Wilesmith JW, Wells GA, Cranwell MP, Ryan JB. 1988. Bovine spongiform encephalopathy: epidemiological studies. Vet. Rec. 123:638–644.
28. World Organisation for Animal Health 2011. Terrestrial Animal Health Code, p 571–588 World Organisation for Animal Health, Paris, France.
SUMBER:
Claudia Guldimann, Michaela Gsponer, Cord Drögemüller, Anna Oevermann, and Torsten Seuberlich. Atypical H-Type Bovine Spongiform Encephalopathy in a Cow Born after the Reinforced Feed Ban on Meat-and-Bone Meal in Europe. J Clin Microbiol. 2012 Dec; 50(12): 4171–4174. doi: 10.1128/JCM.02178-12.
No comments:
Post a Comment